Light concepts for apple orchards
 Print
Print Email
EmailA review of concepts informing orchard practices.
Light interception is one of the most essential considerations for a productive orchard. While this is unlikely to be a new concept, it can be helpful to revisit the impacts and underlying mechanisms of light interception. The following is a review of some light concepts as they relate to orchard practices.
The result of increased light interception has a two-pronged effect:
- Light drives photosynthesis, leading to the fixation of carbon. This carbon is used to produce energy and carbohydrate, which are in turn used to produce dry matter (i.e., the potential for greater biomass in vegetative, woody and fruit tissues).
- Light also acts as a signal, affecting many processes in the plant. It influences the partitioning of resources into fruit versus vegetative tissues, development of anthocyanins, and other processes.
Measuring light quantity and quality
Visible light (400-700 nanometers) is the most important part of the light spectrum for plants. This is because photosynthesis absorbs light in the visible spectrum, and it is by far the most important process in plants influenced by light. Light in this range is measured as photosynthetically active radiation (PAR) or photosynthetic photon flux (PPF). Both indicate the number of photons or light particles.
To help us quantity the energy from light that is available to plants, we can calculate a rate for the number of photons over time. For PAR, we calculate the number of photons with the unit of measure, micromoles (μmol), over a specified area (m-2) and time (s-1).
For example, on a clear, sunny day at noon, PAR is quantified as about 2,000 μmol m-2s-1. On a cloudy day, PAR may be about 400 μmol m-2s-1. For context, apple leaves that develop in full sun can capture light from 900 to 1,000 μmol m-2s-1, which is called the light saturation point. According to Lakso and Seeley, 1978, at this light level, leaf photosynthesis reaches its highest rate, and even though more light is available, leaves cannot capture it.
Available light comes from direct irradiation from the sun as well as diffuse irradiation, or the light scattered by both clouds and by the atmosphere. Diffuse irradiation is deflected in many directions and therefore penetrates the canopy in many locations. Diffuse light is greater and direct light lower in cloudier locations such as in much of Eastern North America.
Light interception and shade
The amount of light intercepted is dependent on characteristics of the canopy including size, density and shading. Smaller canopies intercept more light (Figure 1) as do less dense canopies, which transmit light through the canopy to inner leaves and nearby trees. Shade reduces light interception and can be the result of large or dense canopies. However, even in smaller canopies such as fruiting walls or V-systems, density of the canopy can be too great for good sunlight penetration. This is illustrated well by shade/sunlight patterns in aisleways (Figure 2). The effect of increased shade is less sunlight interception by the shaded leaves and therefore lower rates of photosynthesis (Figure 3).

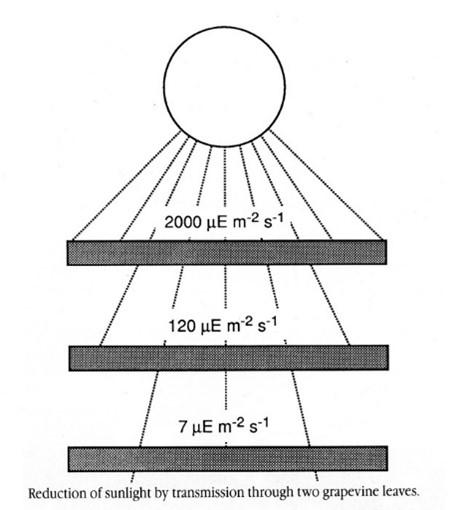
Reducing shade during the emergence and development of individual leaves is important. Leaves which develop in full sun can capture more light and have greater maximum photosynthesis ability compared to leaves that develop in shade. This is mostly irreversible once leaves have formed. The result of lower photosynthetic potential is lower potential productivity and less energy going to fruit and shoots.
After the development of leaves, shade continues to play a role in productivity. There is lower photosynthesis in leaves, fruits and wood in shaded areas compared to exposed areas. It is important to note that this is why pruning is essential to minimize shading and enhance light interception in the canopy. This is especially important in high density planting systems to maximize light interception, grow leaves in direct sun that have higher photosynthetic potential, and support more energy going to fruit.
Impacts of light interception
Greater light interception results in greater net carbon exchange rate, an indicator of photosynthesis (as described above). It also results in:
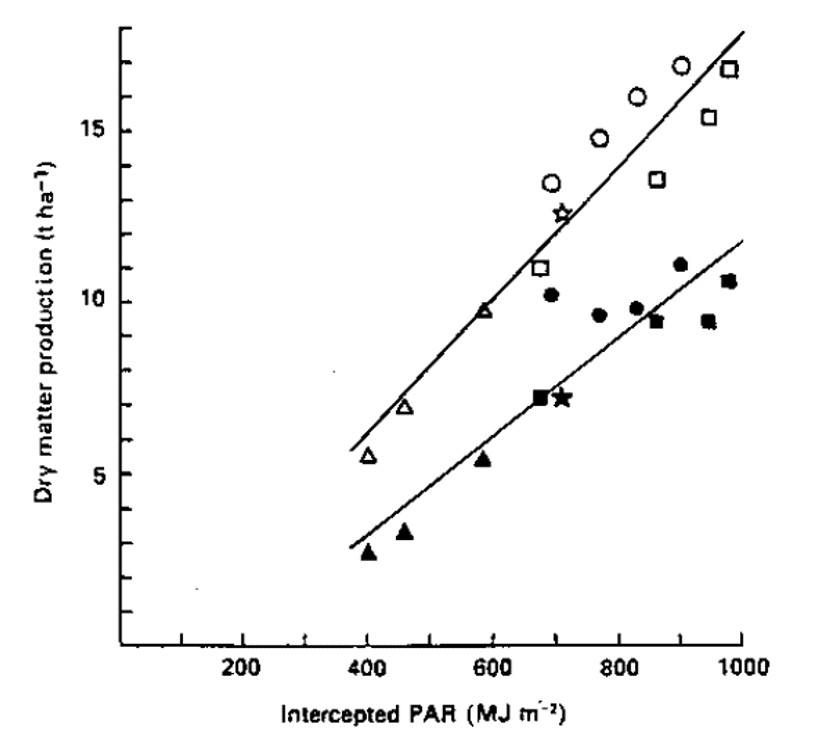
- Increased overall dry matter production (Figure 4).
- Increased fruit set in the current year.
- Flower bud differentiation and development in the following year.
- Improved color and quality.
- Improved wound healing.
Systems with the best sunlight interception
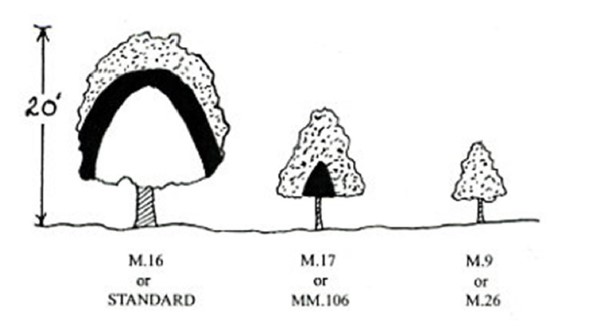
The most light-efficient orchard systems intercept 70% or more of available irradiation. This is important because of the strong relationship between sunlight interception and yield. It is the primary driver of the transition of modern orchard systems to smaller trees with less dense, often two-dimensional canopies.
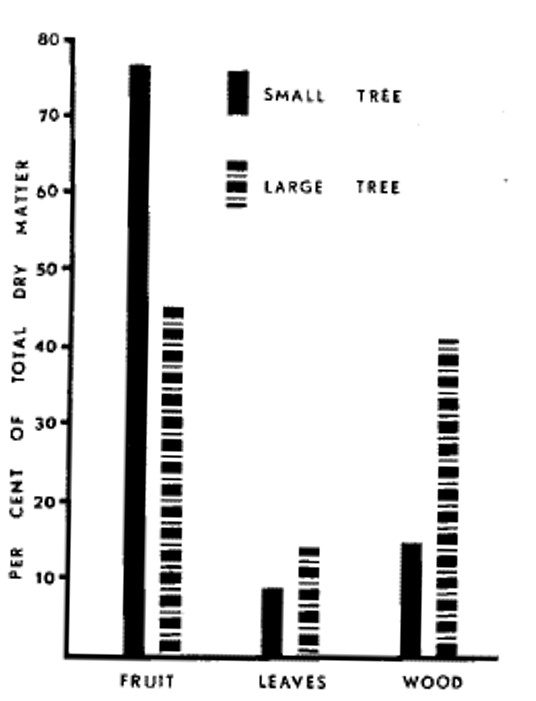
Narrow or shallow canopies (slender spindle, y-trellis, fruiting wall, etc.) have reliably higher sunlight penetration and distribution of light throughout the canopy. Many studies have shown that total yield and yield efficiency (yield per trunk cross sectional area) are higher for canopies with greater sunlight interception and partitioning of dry matter to fruit is greater in smaller trees (Figure 5).
Light and the thinning window
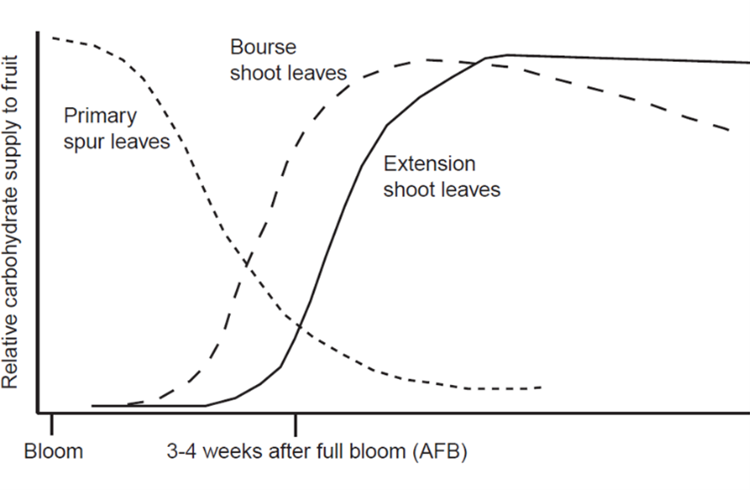
Apple fruit growth is supported by carbohydrate (CHO) partitioning from spur leaves, bourse shoots and extension shoots (Figure 6). Early in the season, primary spur leaves supply CHOs, but this decreases dramatically after bloom (Figure 6, short dashed line). Then, as the bourse shoots (Figure 6, long dashed line) and extension shoots (Figure 6, solid line) grow, they begin to supply CHOs to the fruit. But at approximately three to four weeks after full bloom, there is a much lower CHO supply overall. This timing corresponds to the thinning window when fruitlets are most responsive to thinning applications. Fruitlets compete for CHO while the tree is experiencing a carbohydrate shortage.
In addition, chemical thinners such as NAA suppress carbohydrate production by impacting photosynthetic machinery (photosystem II) in the leaves. All of these factors contribute to fruit thinning and the level of available carbohydrates in the tree indicate why thinning applications are so effective at this time.